Sweet corn is a vegetable and grain dual-use crop with high economic value and industrial advantages. Low temperature stress significantly reduces the germination rate of sweet corn seeds, which has a negative impact on both quality and yield. This study used the chilling sensitive sweet corn inbred line 20hi111 and the chilling tolerant sweet corn inbred line T135 as experimental materials to measure the MDA (malondialdehyde) and H2O2 content, CAT, POD, and SOD enzyme activities, and enzyme gene expression patterns during seed germination under low temperature (10°C) and normal temperature (25°C) treatments. The research results indicated that during low-temperature germination, the H2O2 content and CAT activity of 20hi111 were generally higher than those of T135, while the POD and SOD activities and MDA content were generally lower than those of T135. There was no strict consistency between gene expression and enzyme activity. At low temperature, the expression of ZmCAT1 and ZmCAT3 in 20hi111 was significantly higher than T135, while there was no significant difference in ZmPOD1. In 20hi111, ZmPOD3 was first lower and then higher than T135, and ZmSOD3 and ZmSOD9 were lower than T135 under low temperature treatment. In this study, the activity of antioxidant enzymes and the expression of antioxidant enzyme-related genes in sweet corn inbred lines with different germination characteristics under low temperature were analyzed, which provided some theoretical basis for cultivating sweet corn varieties with low temperature tolerance.
| Published in | Agriculture, Forestry and Fisheries (Volume 13, Issue 6) |
| DOI | 10.11648/j.aff.20241306.11 |
| Page(s) | 224-234 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Sweet Corn, Germination, Low Temperature, Antioxidant Enzyme Activity
Primer name | Primer sequence (5'-3') |
|---|---|
qubi2-F | TGGTTGTGGCTTCGTTGGTT |
qubi2-R | GCTGCAGAAGAGTTTTGGGTACA |
qZmCAT1-F | GAGATCCAAATGGTACGGTATT |
qZmCAT1-R | CAGGCTGTCGTGAGAAGTGC |
qZmCAT3-F | GGGAGAAGGCAACCATAC |
qZmCAT3-R | TGGGTGTCCGAGCGAGT |
qZmPOD1-F | GCCTCCACTTCCACGACT |
qZmPOD1-R | CTTCTTCTCCGACAACCAGC |
qZmPOD3-F | CATCATCGGTGGCACTAAC |
qZmPOD3-R | ACTTTCAGGACCAAGTTTGC |
qZmSOD3-F | TTTGGTTCATTTGAGGCACT |
qZmSOD3-R | CTCCCAGATCAATCCC |
qZmSOD9-F | CGGAGACATCGTGAAATAAA |
qZmSOD9-R | CCACAAAGCACATCGAAAC |
MDA | Malondialdehyde |
CAT | Catalase |
POD | Peroxidase |
SOD | Superoxide Dismutase |
ROS | Reactive Oxygen Species |
RT | Room Temperature Treatment |
LT | Low Temperature Treatment |
qRT-PCR | Quantitative Real-time PCR Analysis |
| [1] | Borowski, A. M., Fritz, V. A., Waters, L. Seed maturity influences of two shrunken-2 sweet germination and vigor corn hybrids. Journal of the American Society for Horticultural Science. 1991, 116(3), 401-404. |
| [2] | Sudjatmiko, S., Chozin, M. Phenological, growth, and yield responses of sweet corn to elevational air temperatures in the humid tropics. Australian Journal of Crop Science. 2024, 18(2), 92-98. |
| [3] | Richard, L., Hassell, Robert, J., Dufault, Tyron, L., Phillips. Low-temperature Germination Response of su, se, and sh2 Sweet Corn Cultivars. Horttechnology. 2003, 13(1), 136-141. |
| [4] | Li, J., Xu, J. G., Lin, C., Guan, Y. J., Hu, J. Effect of priming on germination and physiological characteristics of different types of corn seeds under low-temperature stress. Plant Physiology Journal. 2016, 52(2), 157-166. |
| [5] | Wilson, Franklin, D. Stoichiometry and site specificity of the uncoupling of mitochondrial oxidative phosphorylation by salicylanilide derivatives. Biochemistry. 1969, 8(6), 2475. |
| [6] | Singh, A., Kumar, A., Yadav, S., Singh, I. K. Reactive oxygen species-mediated signaling during abiotic stress. Plant Gene. 2019, 18. |
| [7] | Carocho, M., Ferreira, I. C. A review on antioxidants, prooxidants and related controversy: natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food and Chemical Toxicology. 2013, 51, 15-25. |
| [8] | Wu J. H., Yang L., Sun G. R. Generation of activated oxygen and change of cell defense enzyme activity in leaves of maize seedling under the stress of low temperature. Bulletin of Botanical Research. 2004, 24(4), 456-459. |
| [9] | Zhang, H. Y. Effects of low temperature on seed germination and seedling growth of fresh corn. Plant Physiology Journal. 2013, 49(4), 347-350. |
| [10] | Ramazan, S., Qazi, H. A., Dar, Z. A., John, R. Low temperature elicits differential biochemical and antioxidant responses in maize (Zea mays) genotypes with different susceptibility to low temperature stress. Physiology and molecular biology of plants. 2021, 27(6): 1395-1412. |
| [11] | Wu, Z., Wang, T., Chen, J., Zhang, Y., Lv, G. Sweet corn association panel and genome-wide association analysis reveal loci for chilling-tolerant germination. Scientific reports. 2024, 14(1), 10791. |
| [12] | Jing, Y., Jihua, M., Yongtao, Y. U., Chunyan, L. I., Yongfei, W., Jianguang, H. U. Effects of Chilling on Antioxidant Enzyme Activity and Related Gene Expression Levels During Seed Germination. Journal of Nuclear Agricultural Sciences. 2016. |
| [13] | Souza, D., RS, Vidigal, PS, Scapim, CA, Marques, OJ, Queiroz, DC. Yield and quality of sweet corn at different plant populations. Semin Cienc Agrar. 2013. |
| [14] | Storck, L., Lovato, C., Comassetto, V. Avaliação do rendimento e outras características agronômicas de cultivares de milho doce. Revista Do Centro De Ciências Rurais. 2008, 14(4). |
| [15] | Cutforth, H. W., Shaykewich, C. F. A temperature response function for corn development. Agricultural and Forest Meteorology. 1990, 50(3), 159-171. |
| [16] | Ramazan, S., Jan, N., John, R. Comparative protein analysis of two maize genotypes with contrasting tolerance to low temperature. BMC Plant Biology. 2023, 23(1). |
| [17] | C, K. A., Nalbanto Lu, B. Antifreeze proteins in higher plants. Phytochemistry. 2003, 64(7), 1187-1196. |
| [18] | Xu, P. L., Guo, Y. K., Bai, J. G., Shang, L., Wang, X. J. Effects of long-term chilling on ultrastructure and antioxidant activity in leaves of two cucumber cultivars under low light. Physiologia plantarum. 2008, 132(4), 467-478. |
| [19] | Taşgín, E., Atící, Ö., Nalbantoğlu, B. Effects of salicylic acid and cold on freezing tolerance in winter wheat leaves. Plant Growth Regulation. 2003, 41(3), 231-236. |
| [20] | Janero, D. R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radical Biology & Medicine. 1990, 9(6), 515-540. |
| [21] | Wattanakulpakin, P., Photchanachai, S., Miyagawa, S., Ratanakhanokchai, K. Loss of Maize Seed Vigor as Affected by Biochemical Changes during Hydropriming. Science. 2012, 52(6), 2783. |
| [22] | Li, C. Z., Jiao, J., Wang, G. X. The important roles of reactive oxygen species in the relationship between ethylene and polyamines in leaves of spring wheat seedlings under root osmotic stress. Plant Science. 2004, 166(2), 303-315. |
| [23] | Yu, J. D. Y. S. Physiological basis of different allelopathic reactions of cucumber and figleaf gourd plants to cinnamic acid. Journal of Experimental Botany. 2007, 58(13), 3765-3773. |
| [24] | Río, L. A. D., Corpas, F. J., López-Huertas, E., Palma, J. M. Plant Superoxide Dismutases: Function Under Abiotic Stress Conditions, Antioxidants and Antioxidant Enzymes in Higher Plants. 2018. pp: 1-26. |
| [25] | Guan, L., Scandalios, J. G. Two Structurally Similar Maize Cytosolic Superoxide Dismutase Genes, Sod4 and Sod4A, Respond Differentially to Abscisic Acid and High Osmoticum 1. Plant Physiology. 1998. |
| [26] | Scandalios, J., G. Oxidative stress: molecular perception and transduction of signals triggering antioxidant gene defenses. Brazilian Journal of Medical and Biological Research. 2005, 38(7), 995-1014. |
| [27] | Willekens, H., Inzé, D., Montagu, M. V., Camp, W. V. Catalases in plants. Molecular Breeding. 1995, 1(3), 207-228. |
| [28] | Redinbaugh, M. G., Sabre, M., Scandalios, J. G. The Distribution of Catalase Activity, Isozyme Protein, and Transcript in the Tissues of the Developing Maize Seedling. Plant Physiology. 1990, 92, 375-380. |
| [29] | Puntarulo, S., Sanchez, R. A., Boveris, A. Hydrogen Peroxide Metabolism in Soybean Embryonic Axes at the Onset of Germination. Plant Physiology. 1988, 86(2), 626-630. |
| [30] | Puntarulo, S., Galleano, M., Sanchez, R. A., Boveris, A. Superoxide anion and hydrogen peroxide metabolism in soybean embryonic axes during germination. Biochimica et Biophysica Acta (BBA) - General Subjects. 1991, 1074(2), 277-83. |
| [31] | Bethards, L. A., Skadsen, R. W., Scandalios, J. G. Isolation and characterization of a cDNA clone for the Cat2 gene in maize and its homology with other catalases. Proceedings of the National Academy of Sciences. 1987, 84(19), 6830-6834. |
| [32] | Kuk, Y. I., Shin, J. S., Burgos, N. R., Hwang, T. E., Han, O., Cho, B. H., Jung, S., Guh, J. O. Antioxidative Enzymes Offer Protection from Chilling Damage in Rice Plants. Crop Science. 2003, 43(6). |
| [33] | Hodges, D. M., Andrews, C. J., Johnson, D. A., Hamilton, R. I. Antioxidant enzyme responses to chilling stress in differentially sensitive inbred maize lines. Journal of Experimental Botany. 1997, 5, 5. |
| [34] | Abler, M. L. Isolation and characterization of a genomic sequence encoding the maize Cat3 catalase gene. Plant Molecular Biology. 1993, 22(6), 1031-1038. |
| [35] |
Han-Mei, L., Huai-Yu, Z., Zhen-Bo, T., Yu-Bi, H. Isolation and Expression of the Genes Involved in Tolerance to Low Temperature in Maize. Journal of Maize Sciences. 2007.
http://www.ymkx.com.cn/jms/article/abstract/07020026?st=article_issue |
| [36] | Kratsch, H. A., Wise, R. R. The ultrastructure of chilling stress. Plant Cell & Environment. 2000, 23(4), 337-350. |
| [37] | Miyake, C., Asada, K. Ferredoxin-Dependent Photoreduction of the Monodehydroascorbate Radical in Spinach Thylakoids. Plant & Cell Physiology. 1994, 35(4), 539-549. |
| [38] | Tsang, E. W., Bowler, C., Hérouart, D., Van Camp, W., Villarroel, R., Genetello, C., Van Montagu, M., Inzé, D. Differential regulation of superoxide dismutases in plants exposed to environmental stress. The Plant Cell. 1991, 3(8), 783-92. |
APA Style
Wang, T., Wu, Z., Chen, J., Li, F., Lv, G. (2024). The Effects of Chilling on Antioxidant Enzyme System and Related Gene Expression Levels in Sweet Corn Seeds with Different Germination Characteristics. Agriculture, Forestry and Fisheries, 13(6), 224-234. https://doi.org/10.11648/j.aff.20241306.11
ACS Style
Wang, T.; Wu, Z.; Chen, J.; Li, F.; Lv, G. The Effects of Chilling on Antioxidant Enzyme System and Related Gene Expression Levels in Sweet Corn Seeds with Different Germination Characteristics. Agric. For. Fish. 2024, 13(6), 224-234. doi: 10.11648/j.aff.20241306.11
AMA Style
Wang T, Wu Z, Chen J, Li F, Lv G. The Effects of Chilling on Antioxidant Enzyme System and Related Gene Expression Levels in Sweet Corn Seeds with Different Germination Characteristics. Agric For Fish. 2024;13(6):224-234. doi: 10.11648/j.aff.20241306.11
@article{10.11648/j.aff.20241306.11,
author = {Tingzhen Wang and Zhenxing Wu and Jianjian Chen and Fangjian Li and Guihua Lv},
title = {The Effects of Chilling on Antioxidant Enzyme System and Related Gene Expression Levels in Sweet Corn Seeds with Different Germination Characteristics
},
journal = {Agriculture, Forestry and Fisheries},
volume = {13},
number = {6},
pages = {224-234},
doi = {10.11648/j.aff.20241306.11},
url = {https://doi.org/10.11648/j.aff.20241306.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.aff.20241306.11},
abstract = {Sweet corn is a vegetable and grain dual-use crop with high economic value and industrial advantages. Low temperature stress significantly reduces the germination rate of sweet corn seeds, which has a negative impact on both quality and yield. This study used the chilling sensitive sweet corn inbred line 20hi111 and the chilling tolerant sweet corn inbred line T135 as experimental materials to measure the MDA (malondialdehyde) and H2O2 content, CAT, POD, and SOD enzyme activities, and enzyme gene expression patterns during seed germination under low temperature (10°C) and normal temperature (25°C) treatments. The research results indicated that during low-temperature germination, the H2O2 content and CAT activity of 20hi111 were generally higher than those of T135, while the POD and SOD activities and MDA content were generally lower than those of T135. There was no strict consistency between gene expression and enzyme activity. At low temperature, the expression of ZmCAT1 and ZmCAT3 in 20hi111 was significantly higher than T135, while there was no significant difference in ZmPOD1. In 20hi111, ZmPOD3 was first lower and then higher than T135, and ZmSOD3 and ZmSOD9 were lower than T135 under low temperature treatment. In this study, the activity of antioxidant enzymes and the expression of antioxidant enzyme-related genes in sweet corn inbred lines with different germination characteristics under low temperature were analyzed, which provided some theoretical basis for cultivating sweet corn varieties with low temperature tolerance.
},
year = {2024}
}
TY - JOUR T1 - The Effects of Chilling on Antioxidant Enzyme System and Related Gene Expression Levels in Sweet Corn Seeds with Different Germination Characteristics AU - Tingzhen Wang AU - Zhenxing Wu AU - Jianjian Chen AU - Fangjian Li AU - Guihua Lv Y1 - 2024/11/21 PY - 2024 N1 - https://doi.org/10.11648/j.aff.20241306.11 DO - 10.11648/j.aff.20241306.11 T2 - Agriculture, Forestry and Fisheries JF - Agriculture, Forestry and Fisheries JO - Agriculture, Forestry and Fisheries SP - 224 EP - 234 PB - Science Publishing Group SN - 2328-5648 UR - https://doi.org/10.11648/j.aff.20241306.11 AB - Sweet corn is a vegetable and grain dual-use crop with high economic value and industrial advantages. Low temperature stress significantly reduces the germination rate of sweet corn seeds, which has a negative impact on both quality and yield. This study used the chilling sensitive sweet corn inbred line 20hi111 and the chilling tolerant sweet corn inbred line T135 as experimental materials to measure the MDA (malondialdehyde) and H2O2 content, CAT, POD, and SOD enzyme activities, and enzyme gene expression patterns during seed germination under low temperature (10°C) and normal temperature (25°C) treatments. The research results indicated that during low-temperature germination, the H2O2 content and CAT activity of 20hi111 were generally higher than those of T135, while the POD and SOD activities and MDA content were generally lower than those of T135. There was no strict consistency between gene expression and enzyme activity. At low temperature, the expression of ZmCAT1 and ZmCAT3 in 20hi111 was significantly higher than T135, while there was no significant difference in ZmPOD1. In 20hi111, ZmPOD3 was first lower and then higher than T135, and ZmSOD3 and ZmSOD9 were lower than T135 under low temperature treatment. In this study, the activity of antioxidant enzymes and the expression of antioxidant enzyme-related genes in sweet corn inbred lines with different germination characteristics under low temperature were analyzed, which provided some theoretical basis for cultivating sweet corn varieties with low temperature tolerance. VL - 13 IS - 6 ER -
Institute of Maize and Featured Upland Crops, Zhejiang Academy of Agricultural Sciences, Dongyang, China
Institute of Maize and Featured Upland Crops, Zhejiang Academy of Agricultural Sciences, Dongyang, China
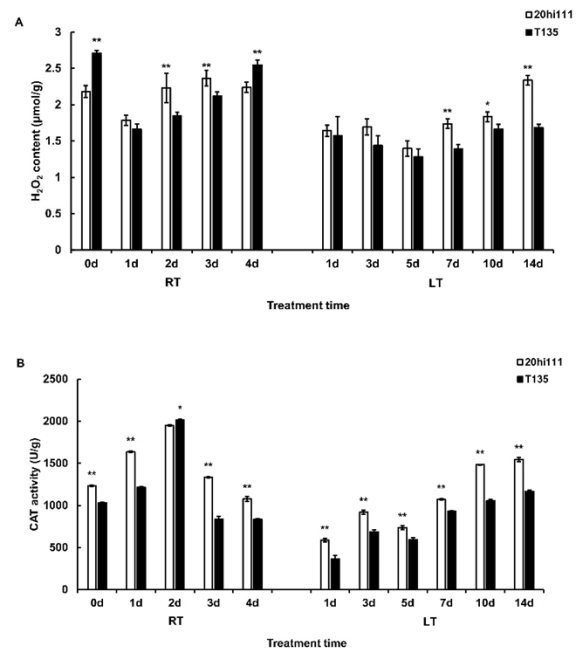
Figure 1. H2O2 content and CAT activity during seed germination of T135 and 20hi111 sweet corn inbred lines under RT and LT. RT: room temperature treatment (25°C). LT: Low temperature treatment (10°C). (A) Determination of H2O2 content. (B) Determination of CAT activity. Standard error is indicated. Asterisks represent Student’s t test significance compared with the control (*P <0.05, **P <0.01).
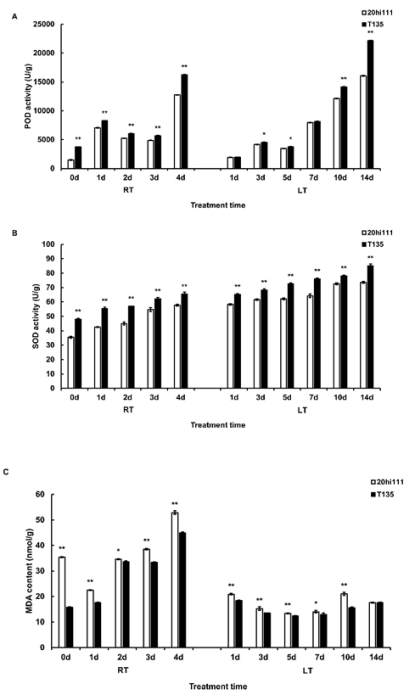
Figure 2. POD, SOD activity and MDA content during seed germination of T135 and 20hi111 sweet corn inbred lines under RT and LT. (A) Determination of POD activity. (B) Determination of SOD activity. (C) Determination of MDA content.
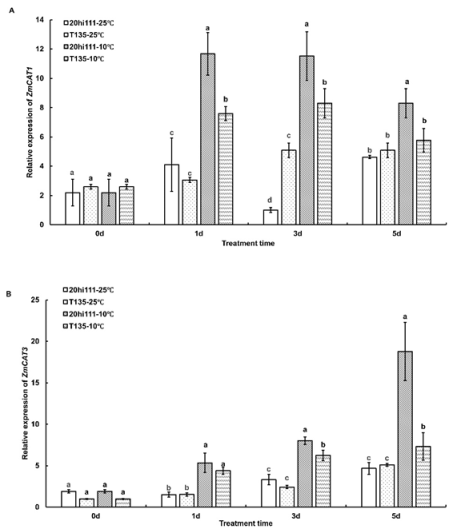
Figure 3. The expression patterns of CAT genes during seed germination of T135 and 20hi111 sweet corn inbred lines under RT (25°C) and LT (10°C). (A) The expression pattern of ZmCAT1. (B) The expression pattern of ZmCAT3. Different letters represent significant differences as determined using one-way ANOVA followed by Duncan’s test (P <0.05).
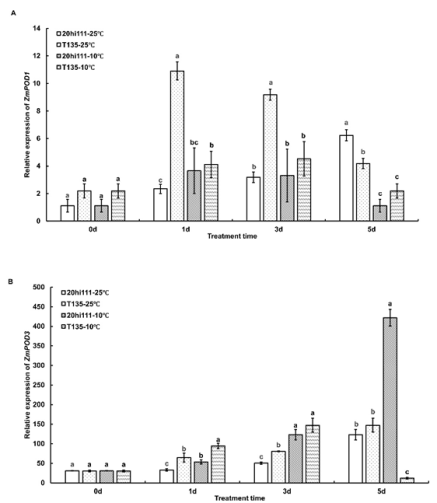
Figure 4. The expression patterns of POD genes during seed germination of T135 and 20hi111 sweet corn inbred lines under RT (25°C) and LT (10°C). (A) The expression pattern of ZmPOD1. (B) The expression pattern of ZmPOD3. Different letters represent significant differences as determined using one-way ANOVA followed by Duncan’s test (P <0.05).
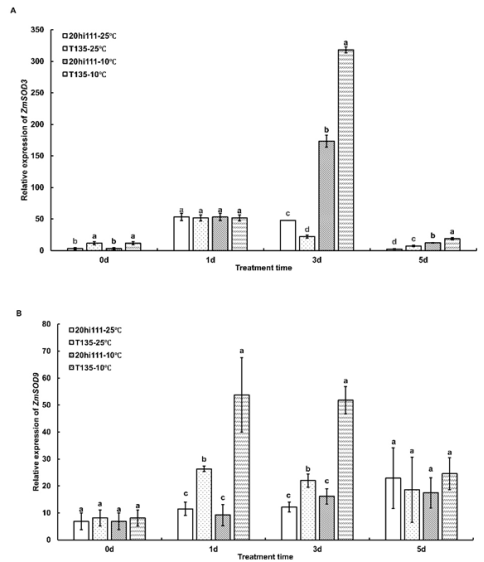
Figure 5. The expression patterns of SOD genes during seed germination of T135 and 20hi111 sweet corn inbred lines under RT (25°C) and LT (10°C). (A) The expression pattern of ZmSOD3. (B) The expression pattern of ZmSOD9. Different letters represent significant differences as determined using one-way ANOVA followed by Duncan’s test (P <0.05).

